Archives
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-11
- 2018-10
- 2018-07
-
Recently the toxicity of TBT was demonstrated at the vascula
2022-02-28
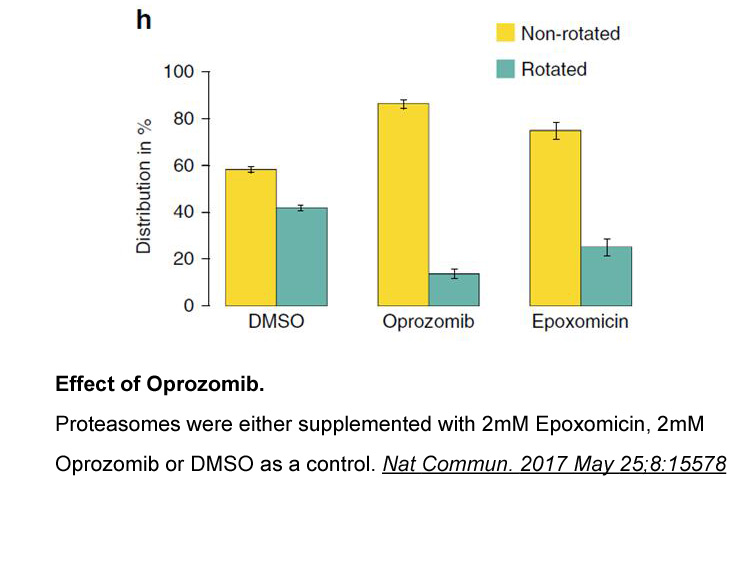
Recently, the toxicity of TBT was demonstrated at the vascular level in animal models (Nath, 2008), and the results obtained so far suggest that TBT may be a potential risk factor for cardiovascular diseases. TBT is capable to alter the coronary vascular reactivity to estradiol (dos Santos et al., 2
-
Here we show that Hippo signaling components are
2022-02-28
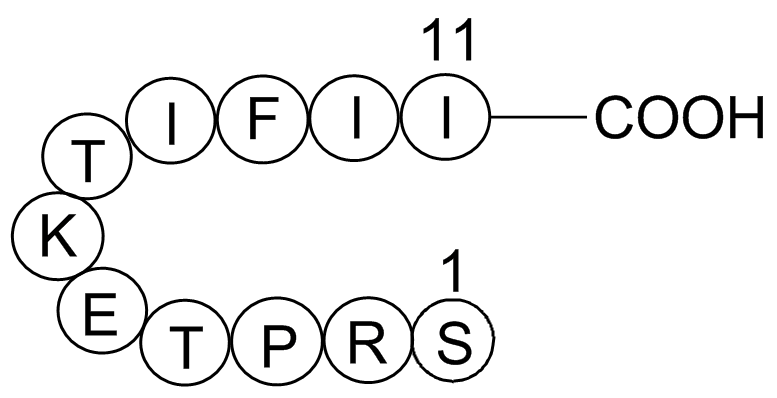
Here, we show that Hippo signaling components are expressed during epicardium formation. To determine the significance of Yap and Taz in the developing epicardium, we generated epicardium-specific Yap/Taz double-knockout mice. Genetic deletion of Yap and Taz using Sema3d mice leads to embryonic leth
-
br Materials and methods br
2022-02-28
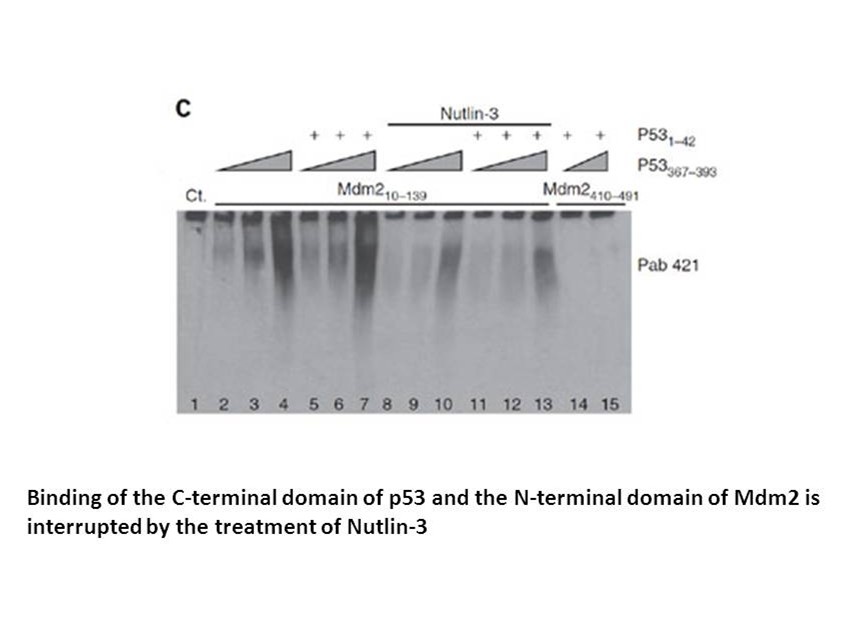
Materials and methods Results Discussion Accurate HER2 assessment for patients with invasive breast cancer is crucial to determining which patients may benefit from HER2-targeted therapy. The most recent ASCO/CAP guidelines have again redefined HER2 gene amplification as determined by dual
-
In conclusion re evaluation of HER
2022-02-28

In conclusion, re-evaluation of HER2 status is necessary to determine the appropriate use of anti-HER2–targeted therapy beyond disease progression. EGFR and c-met amplification, as well as PIK3CA mutation, are rarely associated with acquired resistance. Our results highlight the importance of formal
-
Importantly HDACi mediated effects are cell and
2022-02-28
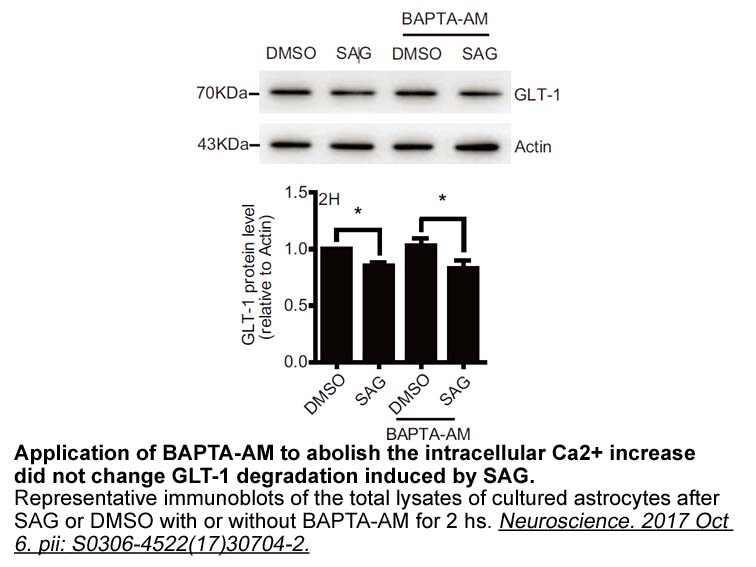
Importantly, HDACi-mediated effects are cell and HDAC specific; different effects and outcomes have been observed in different cell types, targeting different HDAC isoforms. While HDACi exhibit a strong proapoptotic potential in human leukemia M344 19, 20, 21, 22, they have a limited ability to ind
-
br Introduction Marijuana remains the most widely used illeg
2022-02-26
Introduction Marijuana remains the most widely used illegal drug (Murray et al., 2007), and its validated targets include plasma membrane cannabinoid receptors, many of which are found in the central nervous system. The diverse physiological effects produced by marijuana and cannabinoid ligands s
-
In addition to the role
2022-02-26
In addition to the role of TGR5, the FXR-FGF15/19 axis has been implicated in other aspects of glucose metabolism and metabolic disorders. In humans, both obesity and type 2 diabetes mellitus (T2DM) are associated with lower plasma FGF19 levels [67,68]. A derivative of the naturally occurring FXR ag
-
The spatial effects of Ebi
2022-02-26
The spatial effects of Ebi2 involve the creation of a moderate bias in CD4+ T cell distribution toward the LN periphery. Is this modest effect meaningful in an evolutionary sense? In this regard, we think the following considerations are relevant. First, asymmetric CD4+ T cell positioning is highly
-
Generally speaking the regulation of GnIH on gonadotropins s
2022-02-26
Generally speaking, the regulation of GnIH on gonadotropins synthesis and release, although controversial, could be direct and/or indirect via GnRH, among others peptides. In this sense, different scenarios have been reported in vertebrates. In mammalian and avian species, GnIH fibers reach the medi
-
The EEGs of these six patients interestingly
2022-02-26

The EEGs of these six patients interestingly showed focal, multi-focal and generalized spikes, which was an exclusion criteria in our study as we selected our cohort to exclusively represent focal epilepsy. Remarkably all of these cases were diagnosed with paroxysmal exercise-induced dyskinesia (PED
-
br Materials and methods br Results and discussion br Conclu
2022-02-26
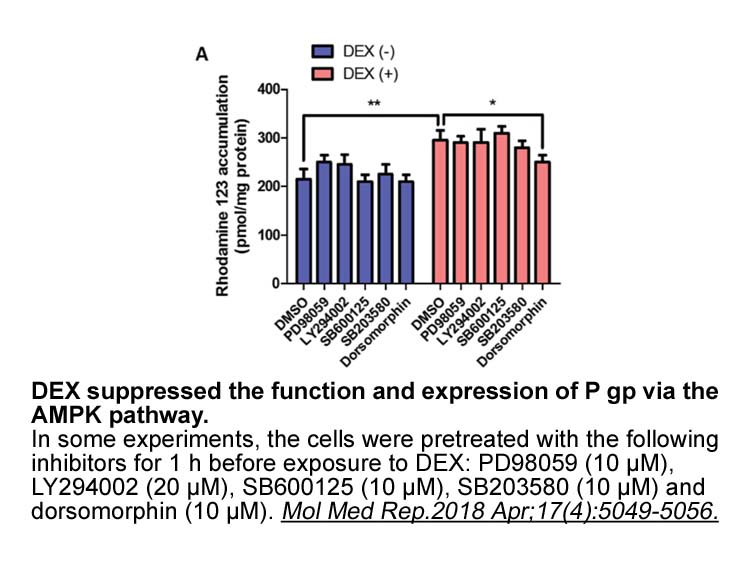
Materials and methods Results and discussion Conclusion Altogether, our data provide an unexpected insight into Cx26 trafficking pathway and gap junction assembly in the cochlea. Since many of the disease-causing mutations in GJB2 impair the trafficking and delivery of Cx26 to the cell surf
-
Developmental studies have reported that some
2022-02-26
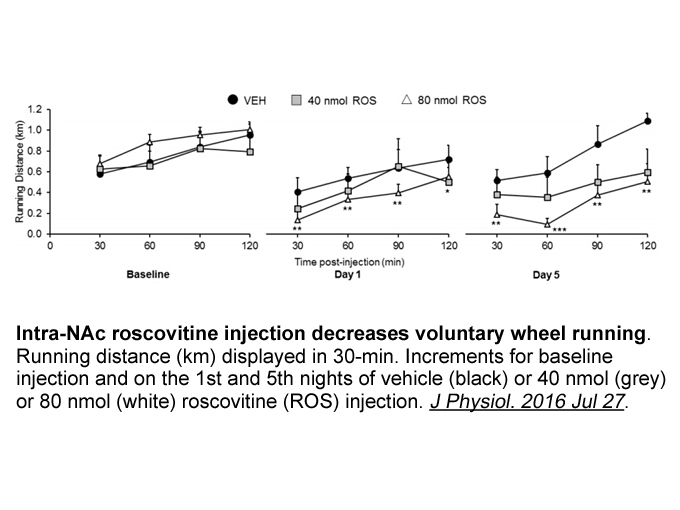
Developmental studies have reported that some subunits of GABAA receptors like α2 and α3 are predominantly expressed during in-utero development, and then substituted for α1 around birth in the rat hippocampus (Brooks-Kayal et al., 2001; Fritschy et al., 1994; Golshani et al., 1997; Kapur and Macdon
-
Rufinamide mg Analysis of published albumin X ray
2022-02-26
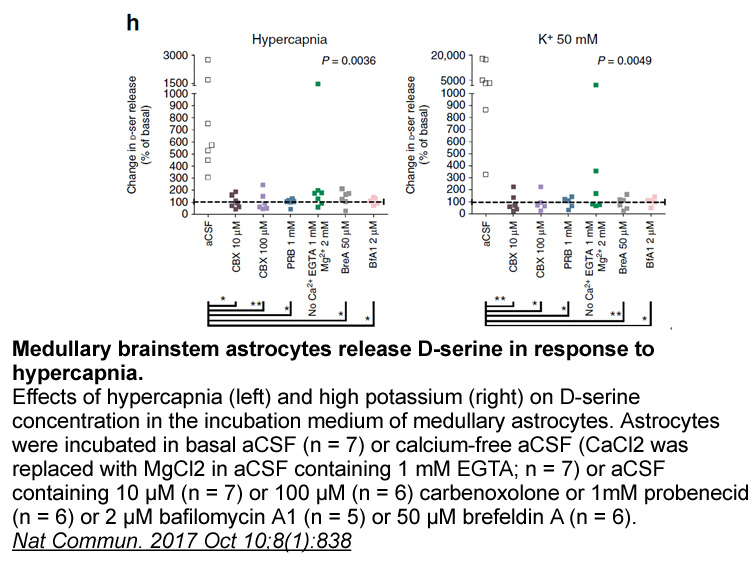
Analysis of published albumin X-ray structures had suggested that an allosteric link may exist, but could not establish whether Zn2+ prevented FFA binding or vice versa. This question was answered through competition experiments monitored by isothermal titration calorimetry (ITC; Fig. 3) for BSA [55
-
The improved glucose tolerance of Ffar R W R
2022-02-26
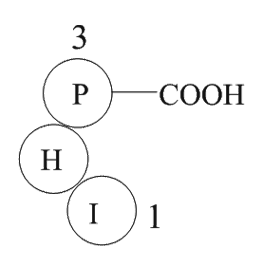
The improved glucose tolerance of Ffar1R258W/R258W mice on HFD was unexpected. In combination with similar fasting blood glucose levels, the significant lower fasting plasma insulin of mutant mice compared to wild-type mice is indicative of improved insulin sensitivity. However, peripheral insulin r
-
The primary cancer lesion was mostly
2022-02-26
The primary cancer lesion was mostly in the uterus and adnexa, followed by the gastrointestinal tract, brain, blood, and kidney and BI 10773 (Fig. 2A). Histopathological findings indicated that most cancers were adenocarcinomas, followed by squamous cell carcinomas, glioblastomas, malignant lymphom
15898 records 474/1060 page Previous Next First page 上5页 471472473474475 下5页 Last page